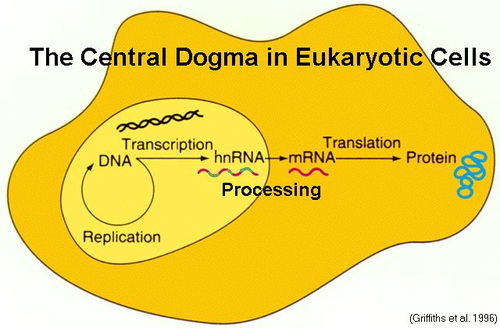
rna加工方法(RNA合成的方法)
1.RNA合成的方法
RNA聚合酶I催化的转录起始RNA聚合酶I催化前rRNA(40S RNA)的合成。前rRNA基因转录起始点上游有两个顺式作用元件(cis acting element),一个是跨越起始点的核心元件(core element),另一个在–100bp处有上游调控元件(upstream control element,UCE)。RNA聚合酶I催化的转录需要2种转录因子,分别称为上游结合因子(upstream binding factor,UBF)和选择性因子1(selective factor1,SL1)。SL1含有4个亚基,一个是TATA盒结合蛋白(TATA-binding protein,TBP),另3个是TBP相关因子(TBP-associated factors,TAF)。UBF与DNA结合令模板DNA发生弯曲,使相距上百bp的UCE和核心元件靠拢,接着SL1和pol I相继结合到UBF-DNA复合物上,完成起始复合物的组建,开始转录
RNA聚合酶Ⅲ催化的转录起始RNA聚合酶Ⅲ催化tRNA,5S rRNA和7S rRNA的转录,tRNA基因的转录初产物是tRNA的前体,经加工后产生多个成熟tRNA。在DNA上的调控序列位于起始转录位点的下游,称为内部启动子。有二个调控区,分别位于编码tRNA D-环和Tψ环的序列,分别称为A盒和B盒。
1.tRNA转录后加工
tRNA的转录后修饰,除了剪接加工外,还包括tRNA链上稀有碱基的形成,以及加上3'端的CCA序列。
2.rRNA的转录后加工
rRNA加工多采用自我剪接的形式。自我剪接的RNA本身形成一种特别的二级结构,称为锤头结构。锤头结构是指复合的茎环组成形态,但其中某些序列上必需是特定的碱基所占据。这种RNA结构,不需要任何蛋白质,就可以水解RNA链上某一特定位点的磷酸二酯键。也就是说,这是一种起催化作用的RNA,现称为核酶。核酶的发现,对酶学、分子生物学,进化生物学都是重要的理论更新,而且,医学上已开始利用人工设计的核酶,去消灭一些作为病原体的RNA病毒或消除一些不利于生命活动的细胞内RNA。
2.mRNA前体的加工过程有哪些步骤
mRNA前体的加工过程有哪些步骤:
原核生物转录作用生成的mRNA属于多顺反子mRNA,即由操纵子机制控制生成的一条mRNA可编码几种不同的蛋白质。原核生物转录生成的初级转录本mRNA不需经过复杂的加工过程即可表现功能,惟一的加工过程是多顺反子mRNA在RnaseⅢ的催化下裂解为单个的顺反子。
真核生物转录生成的是单顺反子mRNA,其前体是非均一RNA(hnRNA)。hnRNA加工过程包括。
剪接
真核生物的基因是一种断裂基因,即其结构基因由若干编码序列和非编码序相间排列而成,其中为蛋白质编码的可转录序列称为外显子,不为蛋白质编码的可转录序列为内含子。转录合成的hnRNA需经过剪接、切掉内含子部分,然后再将外显子部分拼接起来。该过程有多种酶活性物质(包括snRNA)参与。
′末端加“帽”
真核细胞成熟mRNA的5′末端均有一个特殊的结构,即m7Gpp-pmnNp,称为“帽”。帽的生成是在细胞核内进行的,但胞浆中也有酶体系,动物病毒mRNA加帽过程就是在宿主细胞的胞浆内进行的。
′末端加“尾”
mRNA前体分子的3′末端有一段保守序列,由特异的核酸内切酶切去多余的核苷酸,然后在多聚A聚合酶的催化下,由ATP聚合生成多聚A尾。该反应在核内发生,在胞浆中也可继续进行。
碱基修饰
mRNA分子中有少量稀有碱基(如甲基化碱基)是在转录后经化学修饰(如甲基化)而形成的。
选择性加工
某些MRNA前体含有多个3'剪切位点和多聚腺苷酸化位点,因此利用这些选择性位点可产生具有不同3'端非编码区或者具有不同编码能力的RNA产物。通过可变剪接途径可以挑先最保留在MRNA中的外显子,结果单个基因可以合成多种不同的蛋白质。
RNA编辑
在合成并经RNA编辑加工之后,MRNA分子的序列可以发生改变。个别核苷酸可以被置换,添加或者删除。编辑过的MRNA翻译产生了较短脱脂基蛋白B48,由于基缺少一个结合受体的蛋白结构域,因此功能受限。还有好多其他编辑的例子,阵锥虫线粒体MRNA发生RNA编辑,使得最终MRNA中一半以上的尿嘧啶都获自编辑过程。
3.各种RNA转录后加工包括哪些内容
各种RNA转录后加工包括哪些内容
一.mRNA
(1)首尾的修饰
5'端修饰时加m7GpppN帽子结构,在核内完成。
3'端修饰时加上多聚腺苷酸尾巴(polyAAAAAAA),在核内完成。
(2)mRNA的剪接
从DNA模板链转录出的最初转录产物中除去内含子,并将外显子连接起来形成一个连续的RNA分子的过程。
?tp=0_01
(3)mRNA编辑
RNA编辑是指在mRNA水平上改变遗传信息的过程。具体说来,指基因转录产生的mRNA分子中,由于核苷酸的缺失,插入或置换,基因转录物的序列不与基因编码序列互补,使翻译生成的蛋白质的氨基酸组成,不同于基因序列中的编码信息现象。
二.tRNA
(1)5'端前导序列由RNaseP切除。
(2)3'端由tRNA核苷酸转移酶加入CCA-OH作为末端。
(3)还包括稀有碱基生成,甲基化等。
三.rRNA
(1)真核生物核内是45S的转录产物,是三种rRNA的前身。
(2)形成小亚基18S-RNA。
(3)形成大亚基28S,5.8S的rRNA.
视情况掌握把,重点是mRNA转录后的加工修饰。也不知道你想问的是不是这个。希望帮到你!
4.提取制备RNA常用的方法有哪几种,各有何优缺点
常见的RNA提取方法有苯酚法、阴离子去污剂法、LiCl一尿素法、改良的Gomez法、异硫氢酸胍法、CTAB法、热硼酸法及改良热硼酸法、TRIZOL试剂快速提取法。
RNA提取时,就变性剂的选择来说,CTAB(CTAB法)比异硫氰酸胍(RIZOL试剂法)和 SDS(改良热硼酸法)更有效;就CTAB法来说,由于以CTAB为变性剂,同时加入PVP和β一巯基乙醇共同作用变性蛋白、抑制RNase的活性,使用无水乙醇或异丙醇沉淀杂蛋白和总核酸等,然后再选择性地分离出RNA。
5.关于生物化学,问3种RNA如何进行转录加工,加工到哪些地方
真核生物转录起始十分复杂,往往需要多种蛋白因子的协助,转录因子与RNA聚合酶Ⅱ形成转录起始复合体,共同参与转录起始的过程。
根据转录因子的作用特点可分为二类;第一类为普遍转录因子,它们与RNA聚合酶Ⅱ共同组成转录起始复合体时,转录才能在正确的位置开始。除TFⅡD以外,还发现TFⅡA,TFⅡB,TFⅡF,TFⅡE,TFⅡH等,它们在转录起始复合体组装的不同阶段起作用。
第二类转录因子为组织细胞特异性转录因子,这些TF是在特异的组织细胞或是受到一些类固醇激素\生长因子或其它刺激后,开始表达某些特异蛋白质分子时,才需要的一类转录因子。
6.真核生物RNA转录后加工修饰的方式包括哪些
真核生物 一核糖体RNA:基因拷贝数多,在几十到几千之间。
基因成簇排列在一起,由RNA聚合酶I转录生成一个较长的前体,哺乳动物为45S。核仁是rRNA合成与核糖体亚基生物合成的场所。
RNA酶III等核酸内切酶在加工中起重要作用。5SRNA基因也是成簇排列的,由RNA聚合酶III转录,经加工参与构成大亚基。
核糖体RNA可被甲基化,主要在核苷2'羟基,比原核生物甲基化程度高。多数核糖体RNA没有内含子,有些有内含子但不转录。
二转运RNA:由RNA聚合酶III转录,加工与原核相似,但3'端的CCA都是后加的,还有2'-O-甲基核糖。 三信使RNA:真核生物编码蛋白质的基因以单个基因为转录单位,但有内含子,需切除。
信使RNA的原初转录产物是分子量很大的前体,在核内加工时形成大小不等的中间物,称为核内不均一RNA(hnRNA)。其加工过程包括: 1.5'端加帽子:在转录的早期或转录终止前已经形成。
首先从5'端脱去一个磷酸,再与GTP生成5',5'三磷酸相连的键,最后以S-腺苷甲硫氨酸进行甲基化,形成帽子结构。帽子结构有多种,起识别和稳定作用。
2. 3'端加尾:在核内完成。先由RNA酶III在3'端切断,再由多聚腺苷酸聚合酶加尾。
尾与通过核膜有关,还可防止核酸外切酶降解。 3. 内部甲基化:主要是6-甲基腺嘌呤,在hnRNA中已经存在。
可能对前体的加工起识别作用。 三、RNA的拼接 一转运RNA的拼接:由酶催化,酶识别共同的二级结构,而不是序列。
通常内含子插入到靠近反密码子处,与反密码子配对,取代反密码子环。第一步由内切酶切除插入序列,不需ATP;第二步由RNA连接酶连接,需要ATP。
二四膜虫核糖体RNA的拼接:某些四膜虫26S核糖体RNA基因中有一个内含子,其拼接只需一价和二价阳离子及鸟苷酸或鸟苷存在即可自发进行。其实质是磷酸酯的转移反应,鸟苷酸起辅助因子的作用,提供游离3'羟基。
三信使RNA:真核生物编码蛋白质的核基因的内含子属于第二类内含子,左端为GT,右端为AG。先在左端切开,产生的5'末端与3'端上游形成5',2'-磷酸二酯键,构成套索结构。
然后内含子右端切开,两个外显子连接起来。通过不同的拼接方式,可形成不同的信使RNA。
7.真核生物mRNA加工主要包括哪些步骤
真核生物mRNA加工主要包括哪些步骤
真核生物 一核糖体RNA:基因拷贝数多,在几十到几千之间。基因成簇排列在一起,由RNA聚合酶I转录生成一个较长的前体,哺乳动物为45S。核仁是rRNA合成与核糖体亚基生物合成的场所。RNA酶III等核酸内切酶在加工中起重要作用。5SRNA基因也是成簇排列的,由RNA聚合酶III转录,经加工参与构成大亚基。核糖体RNA可被甲基化,主要在核苷2'羟基,比原核生物甲基化程度高。多数核糖体RNA没有内含子,有些有内含子但不转录。 二转运RNA:由RNA聚合酶III转录,加工与原核相似,但3'端的CCA都是后加的,还有2'-O-甲基核糖。 三信使RNA:真核生物编码蛋白质的基因以单个基因为转录单位,但有内含子,需切除。信使RNA的原初转录产物是分子量很大的前体,在核内加工时形成大小不等的中间物,称为核内不均一RNA(hnRNA)。其加工过程包括: 1.5'端加帽子:在转录的早期或转录终止前已经形成。首先从5'端脱去一个磷酸,再与GTP生成5',5'三磷酸相连的键,最后以S-腺苷甲硫氨酸进行甲基化,形成帽子结构。帽子结构有多种,起识别和稳定作用。 2. 3'端加尾:在核内完成。先由RNA酶III在3'端切断,再由多聚腺苷酸聚合酶加尾。尾与通过核膜有关,还可防止核酸外切酶降解。 3. 内部甲基化:主要是6-甲基腺嘌呤,在hnRNA中已经存在。可能对前体的加工起识别作用。 三、RNA的拼接 一转运RNA的拼接:由酶催化,酶识别共同的二级结构,而不是序列。通常内含子插入到靠近反密码子处,与反密码子配对,取代反密码子环。第一步由内切酶切除插入序列,不需ATP;第二步由RNA连接酶连接,需要ATP。 二四膜虫核糖体RNA的拼接:某些四膜虫26S核糖体RNA基因中有一个内含子,其拼接只需一价和二价阳离子及鸟苷酸或鸟苷存在即可自发进行。其实质是磷酸酯的转移反应,鸟苷酸起辅助因子的作用,提供游离3'羟基。 三信使RNA:真核生物编码蛋白质的核基因的内含子属于第二类内含子,左端为GT,右端为AG。先在左端切开,产生的5'末端与3'端上游形成5',2'-磷酸二酯键,构成套索结构。然后内含子右端切开,两个外显子连接起来。通过不同的拼接方式,可形成不同的信使RNA。